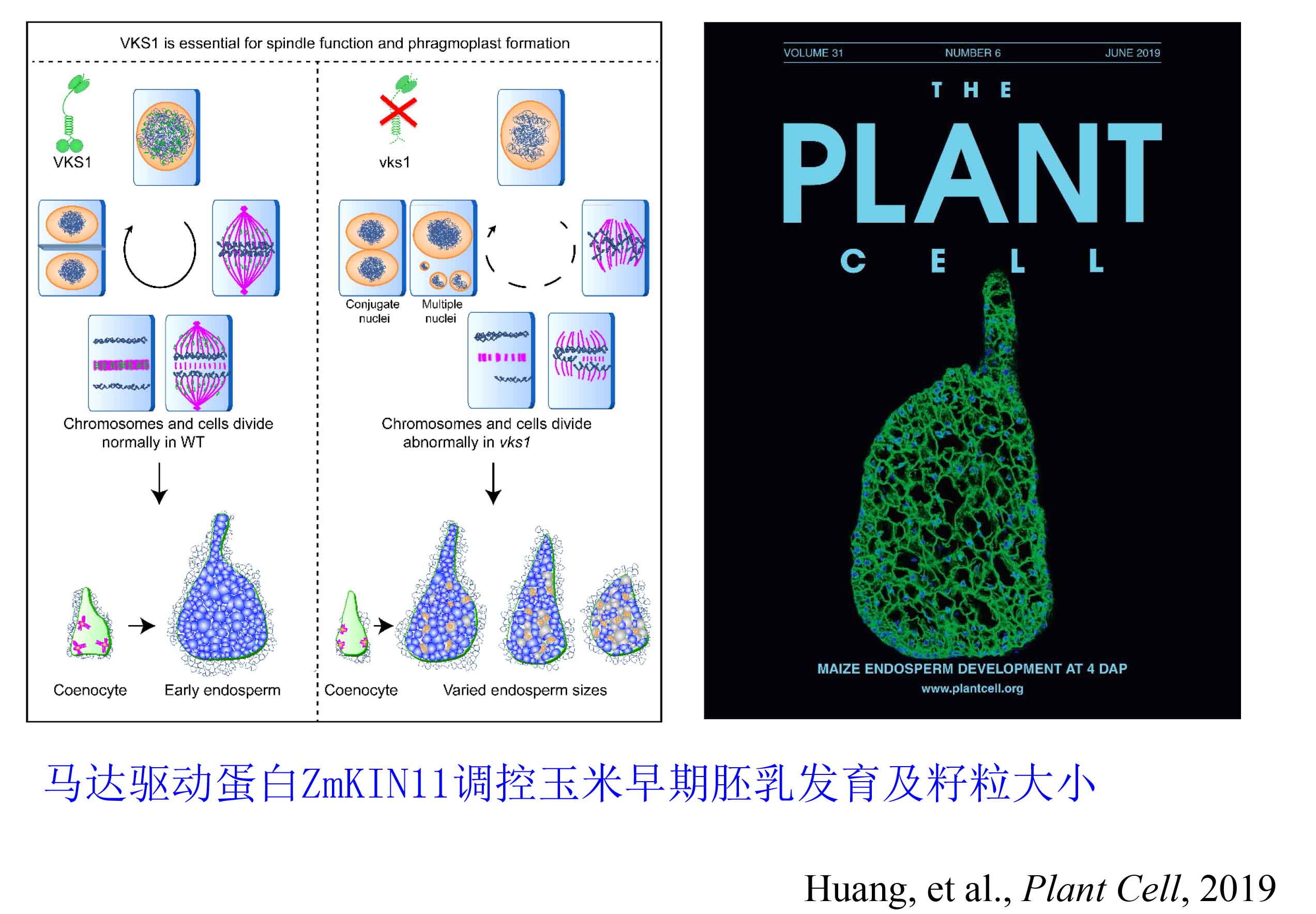
1、 玉米胚乳早期发育机制研究
越来越多证据表明,玉米粒重和品质形成与胚乳早期发育和细胞命运决定密切相关。首先,基因印记调控的基因主要在胚乳中发现,在早期胚乳发育中发挥至关重要作用。很多研究表明籽粒大小受到亲缘效应影响,父本基因的过量表达会导致种子变大,母本基因过量表达会引起种子变小;第二,胚乳体积是由胚乳细胞数目和大小决定的。然而细胞数目在胚乳灌浆前就基本决定了。授粉后8天开始,除胚乳糊粉层和少数亚糊粉层外,整个胚乳细胞已不再分裂。第三,胚乳早期细胞分化形成不同生物学功能区域直接影响胚乳中后期灌浆;第四,胚乳早期发育时间的长短和玉米种子的大小正相关。因此,本实验室针对“玉米胚乳早期发育与细胞命运决定的分子调控网络”开展基础前沿探索研究,阐释胚乳早期发育及特化细胞性状形成的遗传和分子调控机制,为绿色、优质、高产和可持续的玉米遗传改良提供理论基础和模型。
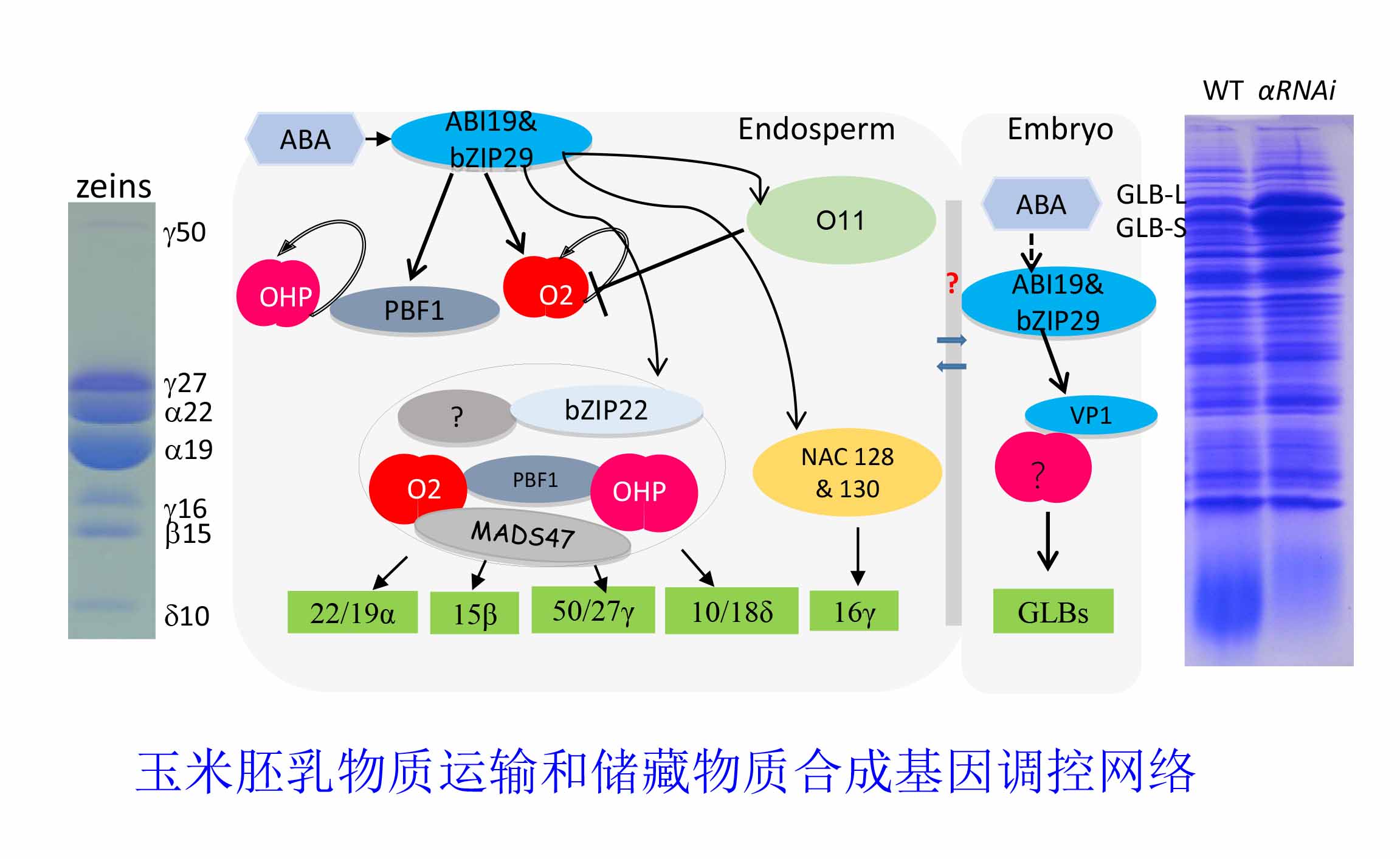
2、玉米胚乳物质运输和储藏物质合成调控网络研究
玉米胚乳中淀粉和储存蛋白合成是一个协同调控过程。蔗糖作为胚乳物质合成的能量来源,其运输进入胚乳的过程也受到了精密的调控,而目前对这方面的研究很少。
本实验室通过遴选关键基因启动子片段筛选胚乳cDNA酵母文库,找到了一些候选转录因子可能参与调控蔗糖运输进入胚乳的过程。另一方面,本实验室通过CRISPR或RNAi转基因和搜索玉米突变体库获得了一些胚乳高表达转录因子的突变体,通过突变体的表型观察和储存物质测定,找到了一些候选突变体影响玉米胚乳淀粉和/或储存蛋白合成。我们还利用EMS诱变创制了优质蛋白玉米突变体库,遴选软质胚乳或灌浆明显不足,但营养和生殖发育没有明显变化的突变体作为重要突变体开展进一步研究,通过MBS辅助克隆基因并进一步研究基因调控机制。通过新调控因子的克隆和各调控因子互作关系的鉴定,构建胚乳物质运输和储存物质合成的调控网络,并找到一些关键基因用于改良底盘品种,获得有自主知识产权更优质高产的玉米。
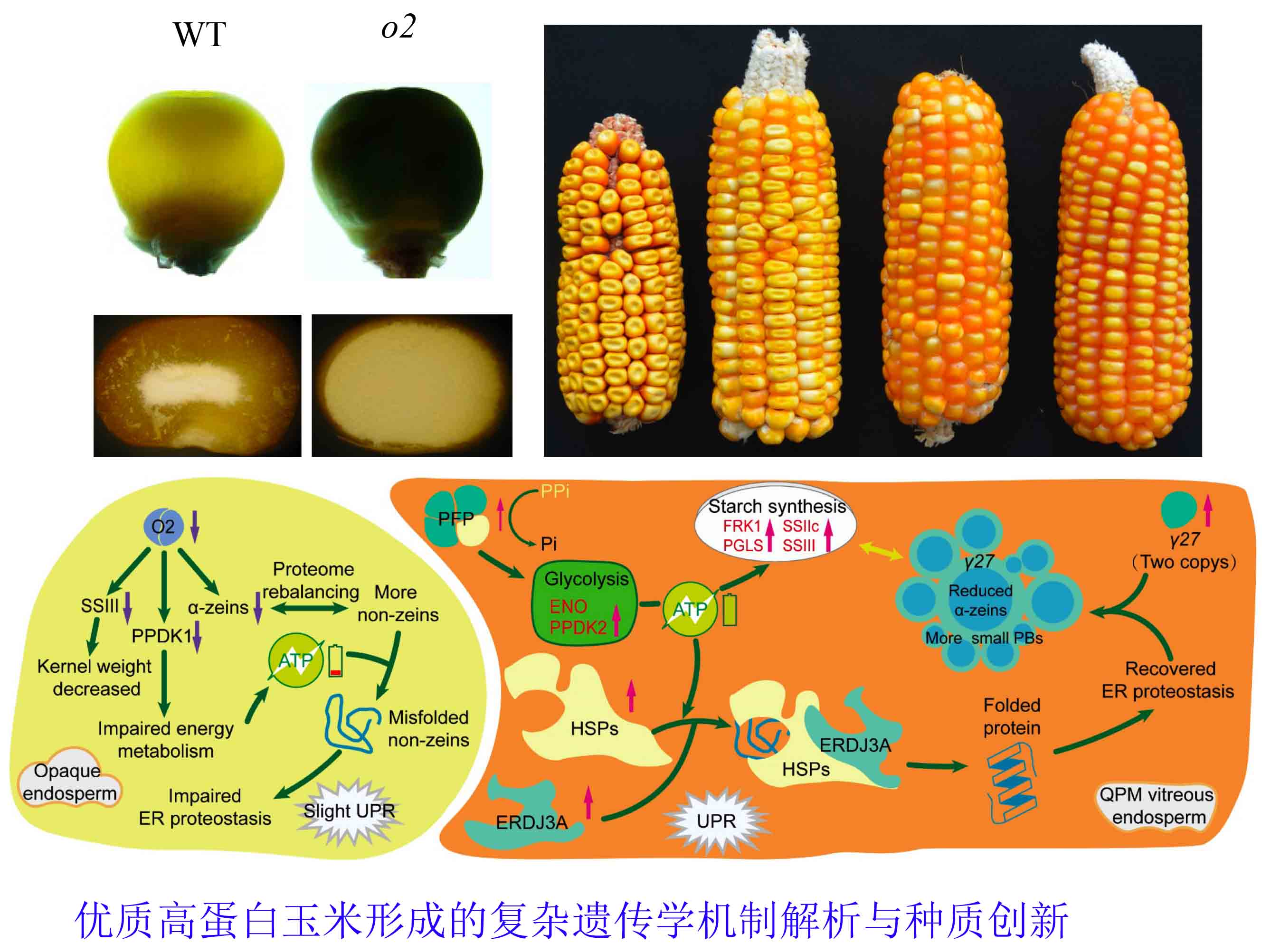
3、优质高蛋白玉米形成的复杂遗传学机制解析与种质创新
新型优质高蛋白玉米分子育种需要三个突破。首先是其胚乳修饰因子数目和机理不清楚,分子标记缺乏,可利用种质资源少。调控27-kD γ-zein的修饰因子(qγ27)已被本实验室克隆(Liu et al., 2016),但自然群体中修饰因子的数目和复杂遗传学基础还未知。本实验室已经完成所有收集材料是否含有修饰因子的评估,发现40%左右的热带自交系在导入α-zein RNAi后能够被完全修饰,而温带只有10%。在今后工作中,本实验室将通过关联、连锁分析挖掘群体中的修饰因子数目,克隆主效QTL并解析它们的功能。
其次是如何克服产量瓶颈。优质蛋白玉米的高赖氨酸品质是由o2突变体提供的,但o2同时造成粒重下降。我们证明了O2不仅调控品质不好的醇溶蛋白表达,也调控主要决定粒重的淀粉合成,所以要使优质蛋白玉米优质却不低产,就必须替换o2寻找新的高赖氨酸供体 (Zhang et al., 2016)。RNAi直接沉默α-zein可以代替o2提高赖氨酸含量而不影响淀粉合成,但用来商业化育种可能会面临转基因限制。由于22-kD 和19-kD α-zein高丰度表达和其拷贝数密切相关,我们可以筛选自然群体中拷贝数少的材料,然后通过遗传重组进一步获得二者拷贝数都少的材料作为高赖氨酸供体。
第三,QPM蛋白品质优越,但蛋白含量偏低。普通玉米自交系总蛋白含量在10-12%,而QPM自交系在8-9%。我们将大刍草(Z. mays ssp. Parviglumis,蛋白含量30%)与B73杂交,然后连续回交构建了B73高蛋白的近等基因系,已经分离到一个主效QTL,它能使B73籽粒总蛋白含量从10.5%提高到14%。我们测量B73-HP的千粒重,发现和B73没有明显区别。从育种角度,这种蛋白含量水平非常理想,如果玉米籽粒总蛋白含量提高至大刍草水平,会影响产量。我们将克隆和功能解析大刍草籽粒高蛋白主效QTL。
本实验室未来五年的目标是聚合高蛋白QTL,低醇溶蛋白拷贝材料和胚乳修饰因子,创制新一代优质高蛋白玉米。